CAPÍTULO 1 Propriedades Gerais das Respostas Imunes
O termo imunidade deriva da palavra latina immunitas, que se refere à proteção contra demandas judiciais que os senadores romanos sofriam durante o seu mandato. Historicamente, a imunidade significava proteção contra doenças e, mais especificamente, contra doenças infecciosas. As células e as moléculas responsáveis pela imunidade constituem o sistema imunológico, e a sua resposta coletiva e coordenada a substâncias estranhas é denominada resposta imunológica.
A função fisiológica do sistema imunológico é a defesa contra micro-organismos infecciosos. Entretanto, até mesmo substâncias estranhas não infecciosas podem desencadear respostas imunológicas. Além disso, os mecanismos que normalmente protegem os indivíduos das infecções e que eliminam as substâncias estranhas também são capazes de provocar lesão tecidual e doença em algumas situações. Portanto, uma definição mais abrangente de resposta imunológica consiste em uma reação a componentes de micro-organismos, bem como a macromoléculas, como proteínas e polissacarídeos, e a pequenas substâncias químicas que são reconhecidas como elementos estranhos, independentemente das consequências fisiológicas ou patológicas dessa reação. Em algumas situações, até mesmo moléculas próprias podem desencadear respostas imunológicas (as denominadas respostas autoimunes). A imunologia é o estudo da resposta imune nessa acepção mais ampla e dos eventos celulares e moleculares que ocorrem após um organismo entrar em contato com micro-organismos e outras macromoléculas estranhas.
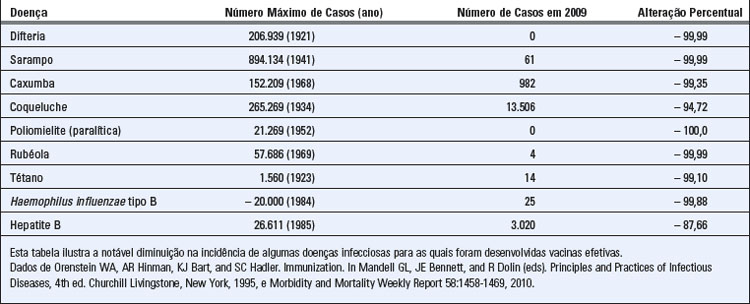
Os historiadores frequentemente atribuem a Tucídides, em Atenas, no século V a.C., o mérito de ter sido o primeiro a usar o termo imunidade contra uma infecção à qual deu o nome de peste (mas que provavelmente não se tratava da peste bubônica que conhecemos hoje em dia). É possível que o conceito de imunidade protetora já existisse muito tempo antes disso, como sugere o antigo costume chinês de tornar as crianças resistentes à varíola através da inalação de um pó feito das lesões cutâneas de pacientes que estavam se recuperando da doença. A imunologia, em sua forma moderna, é uma ciência experimental, na qual as explicações dos fenômenos imunológicos baseiam-se em observações experimentais, com base nas quais são formuladas conclusões. A evolução da imunologia como disciplina experimental depende de nossa habilidade em manipular a função do sistema imunológico em condições controladas. Historicamente, o primeiro exemplo claro dessa manipulação, e que continua constando entre os mais notáveis já registrados, foi a vacinação bem-sucedida, empreendida por Edward Jenner, contra a varíola. Jenner, um médico inglês, observou que mulheres responsáveis pela ordenha das vacas que haviam se recuperado da vaccínia nunca contraíam a varíola, que era mais grave. Com base nessa observação, ele injetou o material de uma pústula de vaccínia no braço de um menino de 8 anos de idade. Quando, posteriormente, esse garoto recebeu inóculo intencional de varíola, ele não desenvolveu a doença. O tratado pioneiro de Jenner sobre a vacinação (do latim vaccinus, relativo a, ou derivado de vacas) foi publicado em 1798. Isso levou a ampla aceitação desse método para induzir imunidade a doenças infecciosas, e a vacinação continua sendo o método mais efetivo de prevenção contra infecções (Tabela 1-1). Um testemunho eloquente da importância da imunologia foi a declaração feita, em 1980, pela Organização Mundial de Saúde, de que a varíola foi a primeira doença a ser erradicada no mundo inteiro por um programa de vacinação.
Desde a década de 1960, houve uma notável transformação na nossa compreensão do sistema imunológico e de suas funções. Os avanços nas técnicas de cultura celular (inclusive a produção de anticorpos monoclonais), na imunoquímica, na metodologia do DNA recombinante e na cristalografia por raios X e a criação de animais geneticamente modificados (particularmente ratos transgênicos e camundongos knockout) fizeram com que a imunologia deixasse de ser uma ciência em grande parte descritiva para se tornar uma ciência na qual diversos fenômenos imunológicos pudessem ser explicados em termos estruturais e bioquímicos. Neste capítulo, vamos descrever as características gerais das respostas imunológicas e apresentar os conceitos que formam as bases da imunologia moderna e que aparecem repetidamente em todo este livro.
IMUNIDADE INATA E ADAPTATIVA
A defesa contra micro-organismos é mediada por reações iniciais da imunidade inata e por respostas tardias da imunidade adaptativa (Fig. 1-1 e Tabela 1-2). A imunidade inata (também denominada imunidade natural ou nativa) proporciona a linha de defesa inicial contra micro-organismos. Consiste em mecanismos de defesa celulares e bioquímicos, que já existem até mesmo antes da infecção e que estão prontos para responder rapidamente a infecções. Esses mecanismos reagem aos micro-organismos e aos produtos das células lesionadas e respondem essencialmente da mesma maneira a infecções repetidas. Os principais componentes do sistema imunológico natural são (1) barreiras físicas e químicas, como os epitélios e as substâncias químicas antimicrobiana produzidas nas superfícies epiteliais; (2) células fagocitárias (neutrófilos, macrófagos), células dendríticas e células assassinas naturais (natural killer − NK); (3) proteínas do sangue, incluindo membros do sistema complemento e outros mediadores da inflamação; e (4) proteínas denominadas citocinas, que regulam e coordenam muitas das atividades das células da imunidade natural. Os mecanismos da imunidade natural são específicos para estruturas que são comuns a grupos de micro-organismos relacionados e podem não distinguir diferenças discretas entre micro-organismos.
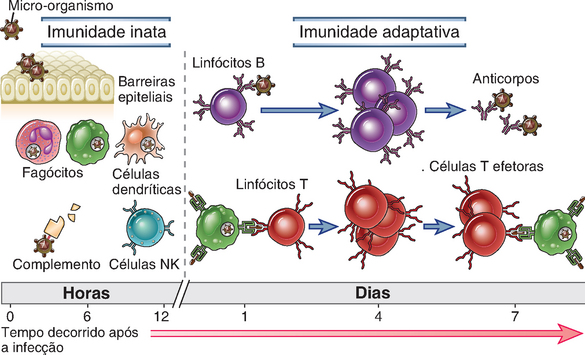
FIGURA 1-1 Imunidade inata e adaptativa. Os mecanismos da imunidade inata fornecem a defesa inicial contra infecções. A resposta imune adaptativa desenvolve-se posteriormente e consiste na ativação dos linfócitos. A cinética das respostas imunológicas inata e adaptativa é uma estimativa e pode variar em diferentes infecções.
TABELA 1-2 Características da Imunidade Inata e Adaptativa
| Inata | Adaptativa | |
|---|---|---|
| Características | ||
| Especificidade | Para moléculas compartilhadas por grupos de micro-organismos relacionados e moléculas produzidas por células do hospedeiro lesionadas | Para antígenos microbianos e não microbianos |
| Diversidade | Limitada; codificada pela linhagem germinativa | Muito grande; são produzidos receptores por recombinação somática de segmentos de genes |
| Memória | Nenhuma | Sim |
| Não reatividade ao próprio | Sim | Sim |
| Componentes | ||
| Barreiras celulares e químicas | Pele, epitélios das mucosas; moléculas antimicrobianas | Linfócitos nos epitélios; anticorpos secretados nas superfícies epiteliais |
| Proteínas do sangue | Complemento, outras | Anticorpos |
| Células | Fagócitos (macrófagos, neutrófilos), células destruidoras naturais | Linfócitos |
Além da imunidade natural, existem outras respostas imunológicas que são estimuladas pela exposição a agentes infecciosos, cuja magnitude e capacidade de defesa aumentam com cada exposição sucessiva a determinado micro-organismo. Como essa forma de imunidade desenvolve-se em resposta à infecção e adapta-se a ela, é denominada imunidade adaptativa ou adquirida. As características que definem a imunidade adaptativa consistem em sua notável especificidade para moléculas distintas e sua capacidade de “lembrar” e responder com mais intensidade em exposições repetidas ao mesmo micro-organismo. O sistema imune adaptativo é capaz de reconhecer e reagir a um grande número de substâncias microbianas e não microbianas. Além disso, possui uma extraordinária capacidade de distinguir micro-organismos e moléculas diferentes e até mesmo micro-organismos e moléculas estreitamente relacionados, motivo pelo qual é também denominado imunidade específica. Algumas vezes, é também designado imunidade adquirida, para ressaltar o fato de que as poderosas respostas protetoras são “adquiridas” por experiência. Os principais componentes da imunidade adquirida consistem em células denominadas linfócitos e seus produtos secretados, tais como os anticorpos. As substâncias estranhas que induzem respostas imunológicas específicas ou que são reconhecidas pelos linfócitos ou por anticorpos são denominadas antígenos.
Os mecanismos de defesa do hospedeiro contra micro-organismos estão presentes, de alguma forma, em todos os seres multicelulares. Esses mecanismos constituem a imunidade natural ou inata. Os mecanismos de defesa mais especializados que constituem a imunidade adquirida são encontrados apenas nos vertebrados. Dois sistemas imunológicos adquiridos funcionalmente similares, porém distintos em nível molecular, desenvolveram-se em diferentes momentos da evolução. Há cerca de 500 milhões de anos, peixes sem maxilas, como a lampreia e a feiticeira, desenvolveram um sistema imunológico singular, contendo diversas células semelhantes aos linfócitos de espécies mais avançadas, capazes de atuar como tal e que até mesmo respondiam à imunização. Os receptores de antígenos nessas células eram receptores variáveis ricos em leucinas, capazes de reconhecer muitos antígenos, mas que eram distintos dos anticorpos e dos receptores de células T que surgiriam mais tarde durante a evolução. Os componentes da imunidade adquirida, inclusive linfócitos com receptores de antígenos altamente diversos, anticorpos e tecidos linfoides especializados, evoluíram, em sua maioria, de modo coordenado em um curto intervalo de tempo nos vertebrados com maxilas (p. ex., tubarões), há cerca de 360 milhões de anos. O sistema imunológico tornou-se também cada vez mais especializado com a evolução.
As respostas imunes natural e adquirida são componentes de um sistema integrado de defesa do hospedeiro, no qual numerosas células e moléculas atuam em cooperação. Os mecanismos da imunidade natural proporcionam uma defesa inicial efetiva contra as infecções. Entretanto, muitos micro-organismos patogênicos evoluíram, tornando-se resistentes à imunidade natural, de modo que a sua eliminação exige a atuação dos mecanismos mais poderosos da imunidade adquirida. Existem muitas conexões entre a imunidade natural e a adquirida. A resposta imune natural aos micro-organismos estimula respostas imunes adquiridas e influi na natureza das mesmas. Em contrapartida, as respostas imunes adaptativas frequentemente atuam ao intensificar os mecanismos protetores da imunidade natural, tornando-os capazes de combater com maior eficácia os micro-organismos patogênicos.
TIPOS DE RESPOSTA IMUNE ADAPTATIVA
Existem dois tipos de resposta imune adaptativa, denominados imunidade humoral e imunidade celular, as quais são mediadas por diferentes componentes do sistema imunológico e cuja função é eliminar diferentes tipos de micro-organismos (Fig. 1-2). A imunidade humoral é mediada por moléculas no sangue e nas secreções das mucosas, denominadas anticorpos, que são produzidos por células denominadas linfócito B (também conhecidas como células B). Os anticorpos reconhecem antígenos microbianos, neutralizam a capacidade dos micro-organismos de infectar e promovem a sua eliminação através de diversos mecanismos efetores. A imunidade humoral é o principal mecanismo de defesa contra micro-organismos extracelulares e suas toxinas, visto que os anticorpos secretados podem ligar-se a esses micro-organismos e toxinas e ajudar na sua eliminação. Os próprios anticorpos são especializados e podem ativar diferentes mecanismos efetores. Por exemplo, diferentes tipos de anticorpos promovem a ingestão de micro-organismos por células do hospedeiro (fagocitose), ligam-se a células de defesa e desencadeiam a liberação de mediadores inflamatórios por essas células, e são ativamente transportados para o lúmen de órgãos que possuem mucosa e através da placenta para fornecer uma defesa contra micro-organismos ingeridos ou inalados e contra infecções do recém-nascido, respectivamente. A imunidade celular, também denominada imunidade mediada por células, é mediada pelos linfócitos T (também designados como células T). Os micro-organismos intracelulares, como os vírus e algumas bactérias, sobrevivem e proliferam no interior dos fagócitos e de outras células do hospedeiro, onde são inacessíveis aos anticorpos circulantes. A defesa contra essas infecções constitui uma função da imunidade celular, que promove a destruição dos micro-organismos que residem nos macrófagos ou a destruição das células infectadas para eliminar os reservatórios da infecção.
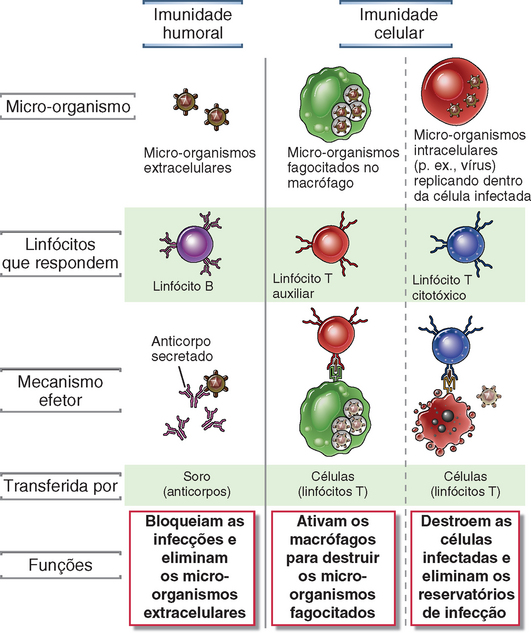
FIGURA 1-2 Tipos de imunidade adaptativa. Na imunidade humoral, os linfócitos B secretam anticorpos que impedem as infecções por micro-organismos extracelulares e os eliminam. Na imunidade celular, os linfócitos T auxiliares ativam os macrófagos para destruir os micro-organismos fagocitados, ou os linfócitos T citotóxicos destroem diretamente as células infectadas.
A imunidade protetora contra um micro-organismo geralmente é induzida pela resposta do hospedeiro ao micro-organismo (Fig. 1-3). A forma de imunidade que é induzida pela exposição a um antígeno é denominada imunidade ativa, visto que o indivíduo imunizado desempenha um papel ativo na resposta ao antígeno. Os indivíduos e os linfócitos que não tiveram exposição a determinado antígeno são denominados virgens (do inglês, naïve), o que significa que são imunologicamente inexperientes. Os indivíduos que já responderam a um antígeno microbiano e que estão protegidos contra exposições subsequentes àquele micro-organismo específico são considerados imunes.
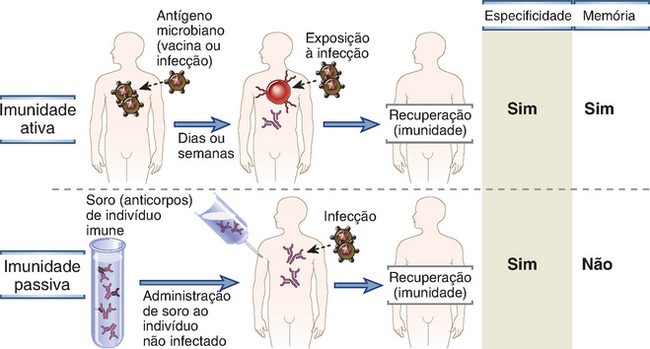
FIGURA 1-3 Imunidade ativa e passiva. A imunidade ativa é conferida pela resposta do hospedeiro a um micro-organismo ou a um antígeno microbiano, enquanto a imunidade passiva é conferida pela transferência adotiva de anticorpos ou de linfócitos T específicos para o micro-organismo. Ambas as formas de imunidade fornecem resistência à infecção e são específicas para antígenos microbianos, mas apenas as respostas imunológicas ativas geram uma memória imunológica. As transferências de células só podem ser realizadas entre doador e receptor geneticamente idênticos (p. ex., camundongos isogênicos) para evitar rejeição das células transferidas.
A imunidade também pode ser conferida a um indivíduo pela transferência de soro ou de linfócitos de um indivíduo especificamente imunizado, constituindo um processo conhecido, em situações experimentais, como transferência adotiva (Fig. 1-3). O receptor dessa transferência torna-se imune ao antígeno específico sem nunca ter sido exposto ou ter respondido a ele. Por conseguinte, essa forma de imunidade é denominada imunidade passiva. A imunidade passiva constitui um método útil para se conferir resistência rapidamente sem a necessidade de esperar o desenvolvimento de uma resposta imunológica ativa. Um exemplo fisiologicamente importante de imunidade passiva é a transferência de anticorpos maternos para o feto, o que permite ao recém-nascido combater as infecções antes que adquira a capacidade de produzir anticorpos. A imunização passiva contra toxinas pela administração de anticorpos de animais imunizados constitui um tratamento que pode salvar a vida no caso de infecções potencialmente letais, como o tétano, e picadas de cobras. A técnica de transferência adotiva também possibilitou a identificação das várias células e moléculas que são responsáveis por mediar a imunidade específica. De fato, a imunidade humoral foi originalmente definida como um tipo de imunidade passível de ser transferida a indivíduos não imunes, ou virgens, através de porções do sangue isentas de células e contendo anticorpos (i.e., plasma ou soro), obtidas de indivíduos previamente imunizados. De modo semelhante, a imunidade celular foi definida como a forma de imunidade que pode ser transferida a animais não imunes por meio de células (linfócitos T) de animais imunizados, mas não por meio do plasma ou do soro.
A primeira demonstração experimental de imunidade humoral foi proporcionada por Emil von Behring e Shibasaburo Kitasato, em 1890. Esses pesquisadores mostraram que, se o plasma de animais que haviam se recuperado de difteria fosse transferido para animais virgens, os animais receptores tornavam-se especificamente resistentes à difteria. Os componentes ativos do soro foram chamados antitoxinas, visto que neutralizam os efeitos patológicos da toxina diftérica. Esse resultado levou ao tratamento da difteria, doença outrora letal, através da administração de antitoxina, uma conquista que foi reconhecida pela concessão do primeiro Prêmio Nobel de Fisiologia ou Medicina a von Behring. No início dos anos 1900, Paul Ehrlich postulou que as células imunes utilizam receptores, aos quais denominou cadeias laterais, para reconhecer toxinas microbianas e secretar subsequentemente esses receptores para combater os micro-organismos. Também criou o termo anticorpos (antikörper, em alemão) para designar as proteínas séricas que se ligam às toxinas, e as substâncias que estimulavam a produção desses anticorpos foram denominadas antígenos. A moderna definição de antígeno abrange substâncias que se ligam a receptores de linfócitos específicos, independentemente de estimularem ou não respostas imunológicas. De acordo com definições estritas, as substâncias que estimulam as respostas imunológicas são denominadas imunógenos. As propriedades dos anticorpos e dos antígenos são descritas no Capítulo 5. Os conceitos de Ehrlich representaram um modelo notavelmente realista da função das células B na imunidade humoral. Essa ênfase inicial nos anticorpos levou a aceitação geral da teoria da imunidade humoral, segundo a qual a defesa do hospedeiro contra infecções é mediada por substâncias presentes nos líquidos corporais (outrora denominados humores).
A teoria da imunidade celular, segundo a qual as células do hospedeiro constituem os principais mediadores da imunidade, foi inicialmente defendida por Elie Metchnikoff. Sua demonstração de que os fagócitos cercavam um espinho espetado em uma larva translúcida de estrela-do-mar, publicada em 1883, foi talvez a primeira evidência experimental de que as células respondem a invasores. Ehrlich e Metchnikoff dividiram o Prêmio Nobel, em 1908, em reconhecimento a suas contribuições para o estabelecimento desses princípios fundamentais da imunidade. A observação de Sir Almroth Wright, no início dos anos 1900, de que fatores presentes no soro imune intensificavam a fagocitose de bactérias ao recobri-las, processo conhecido como opsonização, apoiou a crença de que os anticorpos preparavam os micro-organismos para serem ingeridos por fagócitos. Esses “celularistas” pioneiros não foram capazes de provar que a imunidade específica aos micro-organismos podia ser mediada por células. A teoria celular da imunidade tornou-se firmemente estabelecida na década de 1950, quando foi demonstrado que a resistência a uma bactéria intracelular, Listeria monocytogenes, podia ser transferida adotivamente por células, mas não pelo soro. Na atualidade, sabemos que a especificidade da imunidade celular é devida aos linfócitos que, frequentemente, atuam em conjunto com outras células, como os fagócitos, para eliminar os micro-organismos.
Na prática clínica, a imunidade a um micro-organismo ao qual o indivíduo foi anteriormente exposto é medida de modo indireto, através de ensaios para se detectar a presença de produtos da resposta imunológica (como anticorpos séricos específicos contra antígenos microbianos) ou através da administração de substâncias purificadas obtidas do micro-organismo e medição das reações a essas substâncias. A reação a um antígeno microbiano só é detectável nos indivíduos previamente expostos a esse antígeno; diz-se que esses indivíduos estão “sensibilizados” ao antígeno, e a ocorrência da reação indica “sensibilidade”. Apesar de a reação ao antígeno purificado não ter qualquer função protetora, ela significa que o indivíduo sensibilizado é capaz de desencadear uma resposta imunológica protetora contra o micro-organismo.
PRINCIPAIS CARACTERÍSTICAS DA RESPOSTA IMUNE ADAPTATIVA
Todas as respostas imunológicas humorais e celulares a antígenos exógenos apresentam várias propriedades fundamentais que refletem as propriedades dos linfócitos que medeiam essas respostas (Tabela 1-3).
TABELA 1-3 Principais Características da Resposta Imune Adaptativa
| Característica | Significado Funcional |
|---|---|
| Especificidade | Assegura que a resposta imunológica a determinado micro-organismo (ou antígeno não microbiano) seja dirigida contra esse micro-organismo (ou antígeno) |
| Diversidade | Permite ao sistema imunológico responder a uma grande variedade de antígenos |
| Memória | Aumenta a capacidade de combater infecções repetidas pelo menos micro- organismo |
| Expansão clonal | Aumenta o número de linfócitos específicos para determinado antígeno para fazer frente à capacidade replicativa dos micro-organismos |
| Especialização | Gera respostas que são ideais para a defesa contra diferentes tipos de micro-organismos |
| Contração e homeostasia | Permite ao sistema imunológico recuperar-se de uma resposta, de modo que possa responder efetivamente a novos antígenos que encontre |
| Não reatividade ao próprio | Impede a lesão do hospedeiro durante as respostas a antígenos estranhos |
• Especificidade e diversidade. As respostas imunológicas são específicas para diferentes antígenos e, na verdade, para diferentes porções de uma única proteína complexa, polissacarídio ou outra macromolécula (Fig. 1-4). As partes desses antígenos que são reconhecidas especificamente pelos linfócitos são denominadas determinantes antigênicos ou epítopos. Essa especificidade apurada ocorre porque os linfócitos expressam receptores de membrana que são capazes de distinguir diferenças sutis na estrutura de diferentes epítopos. Os indivíduos não imunes apresentam clones de linfócitos com diferentes especificidades, que são capazes de reconhecer e responder a antígenos estranhos. Esse conceito constitui o princípio básico da hipótese de seleção clonal, que será discutida de modo mais detalhado mais adiante neste capítulo.
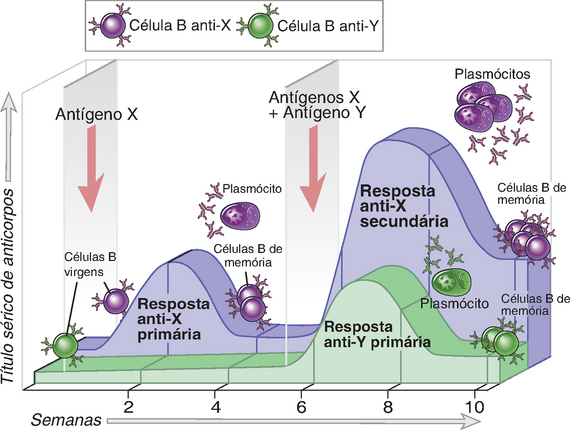
FIGURA 1-4 Especificidade, memória e contração das respostas imunes adaptativas. Os antígenos X e Y induzem a produção de diferentes anticorpos (especificidade). A resposta secundária ao antígeno X é mais rápida e mais vigorosa do que a resposta primária (memória). Os níveis de anticorpos declinam com o passar do tempo após cada imunização (contração, o processo que mantém a homeostasia). As mesmas características são observadas nas respostas imunológicas celulares.
O número total de especificidades antigênicas dos linfócitos de um indivíduo, denominado repertório dos linfócitos, é extremamente grande. Estima-se que o sistema imunológico de uma pessoa tenha a capacidade de discriminar 107 a 109 determinantes antigênicos distintos. Essa capacidade do repertório linfocitário de reconhecer um número muito grande de antígenos, conhecida como diversidade, resulta da variabilidade das estruturas dos sítios de ligação de antígenos presentes nos receptores dos linfócitos. Em outras palavras, existem numerosos clones diferentes de linfócitos, que diferem nas estruturas de seus receptores de antígenos e, portanto, em sua especificidade para antígenos, criando um repertório total que é extremamente diverso. A variação dos receptores de antígenos entre diferentes clones de células T e de células B é o motivo pelo qual esses receptores são descritos como “distribuídos clonalmente”. Os mecanismos moleculares que geram esses receptores de antígenos tão diversos são discutidos no Capítulo 8.
• Memória. A exposição do sistema imunológico a um antígeno estranho aumenta a sua capacidade de responder novamente àquele antígeno específico. As respostas a uma segunda exposição e a exposições subsequentes ao mesmo antígeno, denominadas respostas imunológicas secundárias, geralmente são mais rápidas, de maior intensidade e, com frequência, qualitativamente diferentes da primeira resposta ou resposta imunológica primária ao antígeno (Fig. 1-4). A memória imunológica deve-se ao fato de que cada exposição a um antígeno gera células de memória de vida longa específicas para o antígeno, que são mais numerosas do que as células T virgens específicas para o antígeno, as quais já existem antes da exposição ao antígeno específico. Além disso, essas células de memória possuem características especiais que as tornam mais eficientes na resposta ao antígeno e na eliminação deste do que os linfócitos virgens, que ainda não foram expostos ao antígeno. Por exemplo, os linfócitos B de memória produzem anticorpos que se ligam ao antígeno com maior afinidade do que os anticorpos produzidos nas respostas imunes primárias, e as células T de memória reagem muito mais rapidamente e com mais vigor à estimulação antigênica do que as células T virgens.
• Expansão clonal. Os linfócitos específicos para determinado antígeno sofrem considerável proliferação após a exposição a esse antígeno. A expressão expansão clonal refere-se a um aumento no número de células que expressam receptores idênticos para o mesmo antígeno e que, portanto, pertencem a um clone. Esse aumento nas células específicas para determinado antígeno permite que a resposta imunológica possa fazer frente ao rápido ritmo de divisão dos patógenos.
• Especialização. Conforme já foi mencionado, o sistema imune responde de maneira distinta e especial a diferentes micro-organismos, maximizando a eficiência dos mecanismos de defesa antimicrobianos. Por conseguinte, a imunidade humoral e a imunidade celular são desencadeadas por diferentes classes de micro-organismos ou pelo mesmo micro-organismo em diferentes estágios de infecção (extracelular e intracelular), e cada tipo de resposta imunológica protege o hospedeiro contra aquela classe de micro-organismo. E mesmo no âmbito das respostas imunes humoral e celular, a natureza dos anticorpos ou dos linfócitos T que são gerados pode variar de uma classe de micro-organismo para outra. Abordaremos novamente os mecanismos e o significado funcional dessa especialização nos capítulos posteriores.
• Contração e homeostasia. Todas as respostas imunológicas normais diminuem com o passar do tempo após a estimulação antigênica, de modo que o sistema imunológico retorna a seu estado basal, um estado denominado homeostasia (Fig. 1-4). Essa contração das respostas imunológicas ocorre, em grande parte, pelo fato de que as respostas imunológicas que são desencadeadas por antígenos atuam para eliminá-los, removendo, dessa maneira, um estímulo essencial para a sobrevida e a ativação dos linfócitos. Com exceção das células de memória, os linfócitos que são privados desse estímulo morrem por apoptose.
• Não reatividade ao próprio. Uma das propriedades mais notáveis do sistema imunológico de cada indivíduo normal é a sua capacidade de reconhecer muitos antígenos estranhos (não próprios), de responder a eles e eliminá-los e, ao mesmo tempo, de não reagir de modo prejudicial às substâncias antigênicas próprias do indivíduo. A ausência de resposta imunológica é também denominada tolerância. A tolerância a antígenos próprios, ou autotolerância, é mantida através de vários mecanismos. Esses mecanismos incluem a inativação dos linfócitos que expressam receptores específicos para alguns antígenos próprios, seja eliminando os linfócitos autorreativos, seja suprimindo essas células através das ações de outras células (reguladores). A ocorrência de anormalidades na indução ou na manutenção da autotolerância leva a respostas imunológicas dirigidas contra antígenos próprios (autólogos), o que pode resultar em distúrbios denominados doenças autoimunes. Os mecanismos de autotolerância e as falhas desses mecanismos são discutidos no Capítulo 14.
Essas características da imunidade adquirida são necessárias para que o sistema imune desempenhe a sua função normal de defesa do hospedeiro (Tabela 1-3). A especificidade e a memória permitem ao sistema imune desencadear respostas acentuadas à exposição persistente ou recorrente ao mesmo antígeno e, assim, combater infecções prolongadas ou que ocorrem repetidamente. A diversidade é essencial para que o sistema imunológico possa defender o indivíduo contra os numerosos patógenos potenciais que existem no meio ambiente. A especialização permite que o hospedeiro desenvolva respostas “sob medida” para melhor combater os diferentes tipos de micro-organismos. A contração da resposta permite ao sistema retornar a um estado de repouso após eliminar cada antígeno estranho e a estar preparado para responder a outros antígenos. A autotolerância é essencial para a prevenção de reações prejudiciais contra células e tecidos próprios, mantendo, ao mesmo tempo, um amplo repertório de linfócitos específicos para antígenos estranhos.
As respostas imunológicas são reguladas por um sistema de retroalimentação positiva que amplificam a reação e, por mecanismos de controle, que impedem reações inapropriadas ou patológicas. Quando ativados, os linfócitos desencadeiam mecanismos que aumentam ainda mais a magnitude da resposta. Essa retroalimentação positiva é importante para que o pequeno número de linfócitos específicos contra qualquer micro-organismo possa gerar a resposta necessária para erradicar a infecção. Muitos mecanismos de controle tornam-se ativos nas respostas imunológicas para impedir ativação excessiva dos linfócitos, que poderia causar lesão colateral dos tecidos normais, e para evitar respostas contra antígenos próprios. De fato, todas as respostas imunológicas têm como característica um equilíbrio entre sinais ativadores e inibitórios. Mencionaremos exemplos específicos dessas características fundamentais do sistema imune ao longo deste livro.
COMPONENTES CELULARES DO SISTEMA IMUNOLÓGICO ADAPTATIVO
Os linfócitos, as células apresentadoras de antígenos e as células efetoras são as principais células do sistema imunológico. Os linfócitos são as células que reconhecem e respondem especificamente a antígenos estranhos e que atuam, portanto, como mediadores da imunidade humoral e celular. Existem subpopulações distintas de linfócitos, que diferem em sua maneira de reconhecer os antígenos e nas suas funções (Fig. 1-5). Os linfócitos B são as únicas células capazes de produzir anticorpos. Essas células reconhecem antígenos extracelulares (inclusive de superfície celular) e diferenciam-se em plasmócitos secretores de anticorpos, atuando, assim, como mediadores da imunidade humoral. Os linfócitos T, as células da imunidade celular, reconhecem os antígenos de micro-organismos intracelulares e ajudam os fagócitos a destruí-los ou matam diretamente as células infectadas. As células T não produzem anticorpos. Seus receptores de antígenos são moléculas de membrana distintas dos anticorpos, porém estruturalmente relacionadas a eles (Cap. 7). Os linfócitos T possuem especificidade restrita para os antígenos; eles reconhecem peptídeos derivados de proteínas estranhas que estejam ligadas a proteínas do hospedeiro denominadas moléculas do complexo principal de histocompatibilidade (do inglês, MHC), que são expressas nas superfícies de outras células. Portanto, essas células T reconhecem e respondem a antígenos associados à superfície celular, mas não a antígenos solúveis (Cap. 6). Os linfócitos T consistem em populações funcionalmente distintas, das quais as mais bem definidas são as células T auxiliares (helper) e os linfócitos T citotóxicos (ou citolíticos) (CTL). Em resposta à estimulação antigênica, as células T auxiliares secretam proteínas, denominadas citocinas, que são responsáveis por muitas das repostas celulares da imunidade natural e adquirida e que, portanto, atuam como as “moléculas mensageiras” do sistema imunológico. As citocinas secretadas pelos linfócitos T auxiliares estimulam a proliferação e a diferenciação das próprias células T e ativam outras células, inclusive as células B, os macrófagos e outros leucócitos. Os CTL destroem as células que exibem antígenos estranhos, como as células infectadas por vírus e outros micro-organismos intracelulares. Alguns linfócitos T, denominados células T reguladoras, atuam principalmente para inibir as respostas imunológicas. Uma terceira classe de linfócitos, as células assassinas naturais (natural killer − NK), está envolvida na imunidade natural contra vírus e outros micro-organismos intracelulares. Uma pequena população de linfócitos T que expressam uma proteína de superfície celular encontrada nas células NK é constituída pelas denominadas células NKT; suas especificidades e seu papel na defesa do hospedeiro ainda não estão bem elucidados. Voltaremos a discutir de modo mais detalhado as propriedades dos linfócitos no Capítulo 2, bem como em capítulos subsequentes. As várias classes de linfócitos podem ser diferenciadas pela expressão de proteínas de superfície, que são denominadas moléculas CD e designadas por números (Cap. 2).
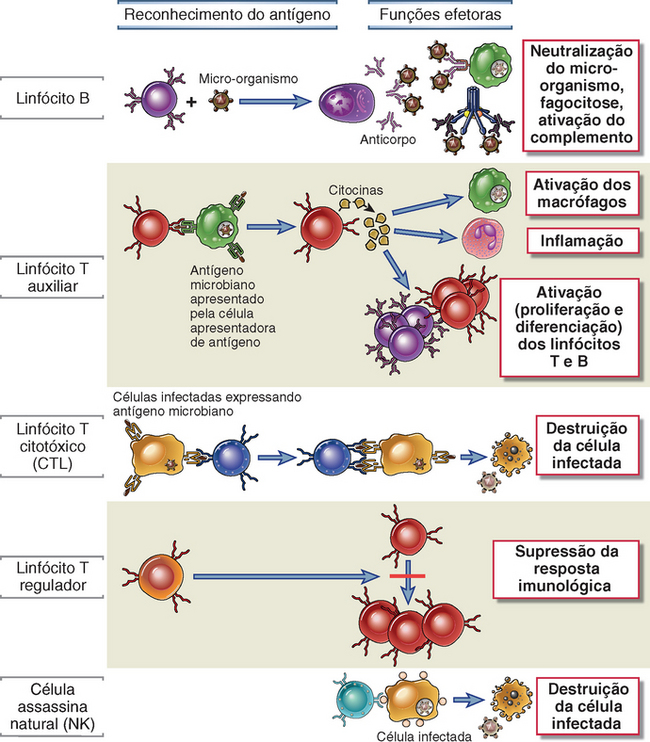
FIGURA 1-5 Classes de linfócitos. Os linfócitos B reconhecem antígenos solúveis e transformam-se em células secretoras de anticorpos. Os linfócitos T auxiliares reconhecem os antígenos presentes na superfície das células apresentadoras de antígenos e secretam citocinas, que estimulam diferentes mecanismos da imunidade e da inflamação. Os linfócitos T citotóxicos reconhecem os antígenos presentes nas células infectadas, destruindo-as. As células T reguladoras suprimem e às vezes até impedem a resposta imunológica (p. ex., a antígenos próprios). As células NK utilizam receptores com diversidade mais limitada do que os receptores de antígenos das células T ou B para reconhecer e destruir seus alvos, tais como as células infectadas.
O início e o desenvolvimento das respostas imunológicas adaptativas exigem que os antígenos sejam capturados e apresentados aos linfócitos específicos. As células que desempenham esse papel são denominadas células apresentadoras de antígenos (do inglês, APC). As APC com maior grau de especialização são as células dendríticas, que capturam os antígenos microbianos provenientes do ambiente externo, transportando-os até os órgãos linfoides e apresentando-os aos linfócitos T virgens, que iniciam as respostas imunológicas. Outros tipos de células atuam como APC em diferentes estágios das respostas celular e humoral. As funções das APC serão descritas no Capítulo 6.
A ativação dos linfócitos pelos antígenos leva à geração de inúmeros mecanismos cuja função é eliminar o antígeno. A eliminação do antígeno frequentemente exige a participação de células denominadas células efetoras, tendo em vista o fato de que medeiam o efeito final da resposta imunológica, que é livrar-se dos micro-organismos. Os linfócitos T ativados, os fagócitos mononucleares e outros leucócitos atuam como células efetoras em diferentes respostas imunológicas.
Os linfócitos e as APC estão concentrados em órgãos linfoides anatomicamente definidos, onde interagem entre si para iniciar as respostas imunológicas. Os linfócitos também estão presentes no sangue; do sangue, podem recircular através dos tecidos linfoides e ser guiados até os locais de exposição antigênica nos tecidos periféricos para eliminar o antígeno específico (Cap. 3).
As células da imunidade natural interagem entre si e com outras células do hospedeiro durante o início e os estágios efetores das respostas imunes naturais e adaptativas. Muitas dessas interações são mediadas por proteínas secretadas, denominadas citocinas. As propriedades e as funções de cada citocina serão descritas quando abordarmos as respostas imunológicas em que essas proteínas desempenham papéis importantes. A seguir, apresentamos um resumo de algumas das características gerais e categorias funcionais das citocinas.
CITOCINAS, MEDIADORES SOLÚVEIS DO SISTEMA IMUNOLÓGICO
As citocinas, que consistem em um grande grupo heterogêneo de proteínas solúveis produzidas por muitos tipos diferentes de células, medeiam e regulam todos os aspectos da imunidade natural e adaptativa. O genoma humano contém cerca de 180 genes que podem codificar proteínas com as características estruturais das citocinas. A nomenclatura das citocinas é um tanto aleatória, e muitas delas são arbitrariamente designadas com base em uma das atividades biológicas descobertas (p. ex., fator de necrose tumoral, interferons), enquanto outras são denominadas interleucinas, com um número como sufixo, visto que se acreditava serem produzidas pelos leucócitos e neles atuarem.
Em geral, as citocinas não são armazenadas como moléculas pré-formadas, e sua síntese é iniciada por nova transcrição gênica como resultado da ativação celular. Essa ativação da transcrição é transitória, e os RNA mensageiros que codificam a maioria das citocinas são instáveis e, com frequência, rapidamente degradados, de modo que a síntese de citocinas também é transitória. Além disso, a produção de algumas citocinas também pode ser regulada por processamento do RNA e por mecanismos pós-tradução, como a liberação proteolítica de um produto ativo de um precursor inativo. Uma vez sintetizadas, as citocinas são rapidamente secretadas, resultando em um surto de liberação, quando necessário.
As citocinas compartilham muitas outras propriedades gerais. Uma citocina pode atuar em diversos tipos de células e exercer múltiplos efeitos biológicos, propriedade designada como pleiotropismo. Por outro lado, várias citocinas podem exercer a mesma ação, e são descritas como redundantes. Uma citocina pode estimular ou inibir a produção de outras, e as citocinas podem antagonizar umas às outras ou produzir efeitos aditivos ou sinérgicos.
As citocinas atuam, em sua maioria, nas proximidades do local em que são produzidas, seja na mesma célula que secreta a citocina (ação autócrina) ou em uma célula adjacente (ação parácrina). As células T frequentemente secretam citocinas no sítio de contato com as APC, constituindo a denominada sinapse imunológica (Cap. 9). Isso pode ser um motivo pelo qual as citocinas frequentemente atuam sobre células que estão em contato com células que as produzem. Quando sintetizadas em grandes quantidades, as citocinas podem entrar na circulação e atuar a distância do seu local de produção (ação endócrina). O fator de necrose tumoral (TNF) é um exemplo de citocina que possui importantes efeitos locais e distantes (sistêmicos).
Algumas citocinas são mediadoras e reguladoras da imunidade natural. São produzidas por células da imunidade natural, como as células dendríticas, os macrófagos e os mastócitos, e impulsionam o processo da inflamação ou contribuem para a defesa contra infecções virais. Outras citocinas, particularmente aquelas produzidas por subgrupos de células T auxiliares, contribuem para a defesa do hospedeiro mediada pela imunidade adaptativa e também regulam as respostas imunológicas. Membros dessa categoria de citocinas também são responsáveis pela ativação e pela diferenciação das células T e das células B. Algumas citocinas são fatores de crescimento para a hematopoiese e regulam a geração de diferentes tipos de células do sistema imune de seus precursores na medula óssea.
Em geral, as citocinas da imunidade inata e da imunidade adquirida são produzidas por diferentes populações de células, atuam sobre células-alvo diferentes e apresentam propriedades distintas. Todavia, essas distinções não são absolutas, visto que a mesma citocina pode ser produzida durante respostas imunológicas naturais e adaptativas, e diferentes citocinas produzidas durante essas respostas podem ter funções sobrepostas.
VISÃO GERAL DAS RESPOSTAS IMUNES AOS MICRO-ORGANISMOS
Agora que já apresentamos os principais componentes do sistema imune e suas propriedades, é conveniente resumir os princípios das respostas imunológicas a diferentes tipos de micro-organismos. Esse resumo irá constituir uma base para os tópicos discutidos em todo o livro. O sistema imune deve combater micro-organismos numerosos e diversos. Como veremos de modo sucinto, as respostas imunes a todos os patógenos compartilham algumas características, e as respostas a diferentes classes desses micro-organismos podem ter características singulares. Os modos pelos quais essas reações imunológicas adquiridas são iniciadas, coordenadas e controladas constituem questões fundamentais da imunologia. Começamos por uma discussão sobre a resposta imune natural.
A Imunidade Inata
O sistema imunológico natural bloqueia a entrada de micro-organismos e elimina ou limita o crescimento de muitos micro-organismos capazes de colonizar os tecidos. Os principais locais de interação entre os indivíduos e seu ambiente — a pele e os tratos gastrointestinal e respiratório — são revestidos por epitélios contínuos, que funcionam como barreiras para impedir a entrada de micro-organismos provenientes do ambiente externo. Se os micro-organismos tiverem sucesso em romper as barreiras epiteliais, deparam-se com as células da imunidade natural. A resposta imune natural celular aos micro-organismos consiste em dois tipos principais de reações — inflamação e defesa antiviral. Inflamação refere-se ao processo de recrutamento de leucócitos e proteínas plasmáticas do sangue, seu acúmulo nos tecidos e sua ativação para destruir os micro-organismos. Muitas dessas reações envolvem citocinas, que são produzidas por células dendríticas, macrófagos e outros tipos de células da imunidade natural. Os principais leucócitos que são recrutados na inflamação são os neutrófilos (cuja sobrevida nos tecidos é de curta duração) e os monócitos (que são transformados em macrófagos teciduais). Esses fagócitos expressam em sua superfície receptores que se ligam aos micro-organismos e os ingerem, bem como outros receptores que reconhecem diferentes moléculas microbianas e ativam as células. Com a ativação desses receptores, os fagócitos produzem radicais reativos de oxigênio e nitrogênio e enzimas lisossômicas, que destroem os micro-organismos que foram ingeridos. Os macrófagos residentes nos tecidos desempenham, em grande parte, as mesmas funções. Defesa antiviral consiste em uma reação mediada por citocinas, em que as células adquirem resistência à infecção viral, e na destruição pelas células NK das células infectadas por vírus.
Os micro-organismos que são capazes de resistir a essas reações de defesa nos tecidos podem entrar no sangue, onde são reconhecidos pelas proteínas circulantes da imunidade natural. Entre as proteínas plasmáticas mais importantes da imunidade natural, encontram-se os componentes da via alternativa do sistema complemento. Quando essa via é ativada por superfícies microbianas, são gerados produtos de clivagem proteolítica, que medeiam as respostas inflamatórias, revestem os micro-organismos para fagocitose intensificada e lisam diretamente os micro-organismos. (Como iremos discutir mais adiante, o complemento também pode ser ativado por anticorpos — é a denominada via clássica, por motivos históricos —, com as mesmas consequências funcionais.) Muitas das proteínas circulantes entram nos locais de infecção durante as respostas inflamatórias e, assim, ajudam a combater os micro-organismos nos tecidos extravasculares.
As reações da imunidade natural são efetivas para controle e até mesmo erradicação das infecções. Entretanto, uma característica essencial de muitos micro-organismos patogênicos é o fato de terem evoluído para resistir à imunidade natural. A defesa contra esses patógenos exige os mecanismos mais poderosos e especializados da imunidade adquirida, que os impedem de invadir e replicarem-se nas células e tecidos do hospedeiro.
A Imunidade Adaptativa
O sistema imune adaptativo utiliza três estratégias principais para combater a maioria dos micro-organismos.
• Os anticorpos secretados ligam-se aos micro-organismos extracelulares, bloqueiam a sua capacidade de infectar células do hospedeiro e promovem sua ingestão e subsequente destruição pelos fagócitos.
• As células T auxiliares aumentam a capacidade microbicida dos fagócitos, que ingerem os micro-organismos e os destroem.
• Os linfócitos T citotóxicos (CTL) destroem as células infectadas por micro-organismos que são inacessíveis aos anticorpos e à destruição fagocítica.
O objetivo da resposta adquirida consiste em ativar um ou mais desses mecanismos de defesa contra micro-organismos diversos que podem estar em diferentes localizações anatômicas, como o lúmen intestinal, a circulação ou dentro de células.
Todas as respostas imunes adaptativas desenvolvem-se por etapas, e cada etapa corresponde a reações específicas dos linfócitos (Fig. 1-6). Começamos essa visão geral da imunidade adquirida pela primeira etapa, que é o reconhecimento dos antígenos.
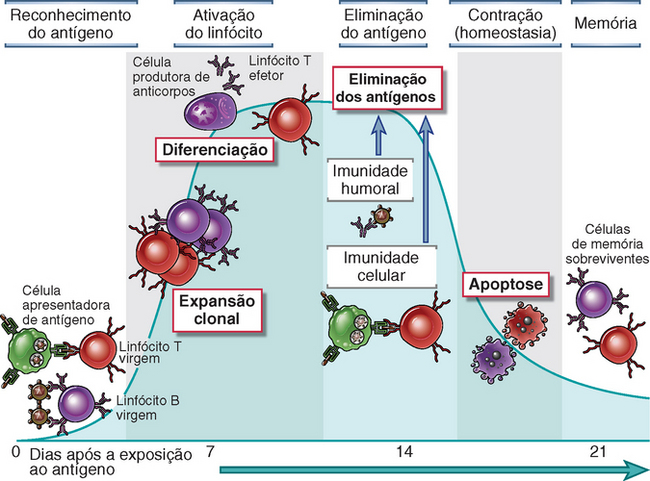
FIGURA 1-6 Fases da resposta imune adaptativa. As respostas imunes adaptativas consistem em fases distintas, das quais as três primeiras são o reconhecimento do antígeno, ativação dos linfócitos e eliminação do antígeno (a fase efetora). A resposta sofre contração (declina) à medida que os linfócitos estimulados pelo antígeno morrem por apoptose, restaurando a homeostasia, e as células específicas para o antígeno em questão que sobrevivem são responsáveis pela memória. A duração de cada fase pode variar em diferentes respostas imunológicas. O eixo y representa uma medida arbitrária da magnitude da resposta. Esses princípios aplicam-se à imunidade humoral (mediada pelos linfócitos B) e à imunidade celular (mediada pelos linfócitos T).
Captura e Apresentação dos Antígenos Microbianos
Como o número de linfócitos virgens específicos para qualquer antígeno é muito pequeno (da ordem de 1 em 105 a 106 linfócitos), e a quantidade do antígeno disponível também pode ser pequena, são necessários mecanismos especiais para capturar os micro-organismos, concentrar seus antígenos na localização correta e apresentar os antígenos a linfócitos específicos. As células dendríticas são as APC que apresentam os peptídeos microbianos aos linfócitos T CD4+ e CD8+ virgens, iniciando respostas imunológicas adquiridas aos antígenos proteicos. As células dendríticas localizadas nos epitélios e nos tecidos conjuntivos capturam os micro-organismos, digerem suas proteínas em peptídeos e expressam, em sua superfície, os peptídeos ligados a moléculas MHC, isto é, as moléculas especializadas de apresentação de peptídeos ao sistema imune adaptativo. As células dendríticas transportam sua carga antigênica até os gânglios satélites e estabelecem residência nas mesmas regiões dos gânglios linfáticos por onde os linfócitos T virgens recirculam continuamente. Dessa maneira, a probabilidade de um linfócito com receptores de antígeno entrar em contato com esse antígeno é acentuadamente aumentada pela concentração do antígeno em uma forma identificável na localização anatômica correta. As células dendríticas também exibem os peptídeos dos micro-organismos em outros tecidos linfoides, como no baço.
Micro-organismos ou antígenos microbianos intactos que entram nos gânglios linfáticos e no baço são reconhecidos em sua forma não processada (nativa) por linfócitos B específicos. Existe também um tipo especial de macrófago, capaz de reter em sua superfície determinados antígenos e “apresentá-los” na sua forma nativa aos linfócitos B.
Reconhecimento dos Antígenos pelos Linfócitos
Existem linfócitos específicos para um grande número de antígenos antes mesmo da exposição ao antígeno, e, quando um antígeno entra no hospedeiro, seleciona as células específicas e as ativa (Fig. 1-7). Esse conceito fundamental é denominado hipótese de seleção clonal. Sugerido pela primeira vez por Niels Jerne, em 1955, esse conceito foi enunciado com mais clareza por Macfarlane Burnet, em 1957, como hipótese para explicar como o sistema imunológico era capaz de responder a um grande número e a uma ampla variedade de antígenos. De acordo com essa hipótese, clones de linfócitos específicos para determinado antígeno desenvolvem-se antes e independentemente de sua exposição ao antígeno. Um “clone” refere-se a um linfócito e sua progênie, de uma única especificidade. Uma característica do sistema imunológico consiste na geração de um número muito grande de clones durante a maturação dos linfócitos, maximizando, assim, o potencial de reconhecimento de diversos micro-organismos.
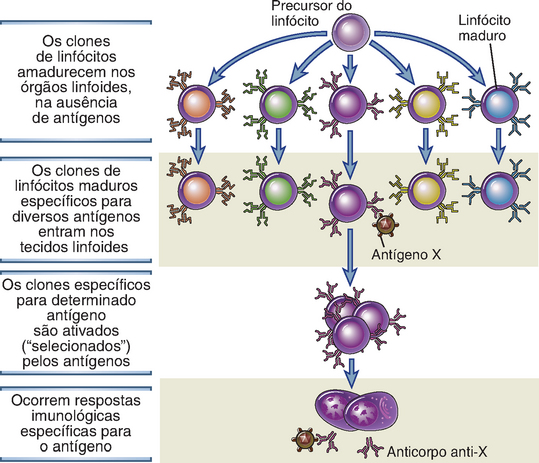
FIGURA 1-7 A hipótese de seleção clonal. Cada antígeno (X ou Y) seleciona um clone preexistente de linfócitos específicos e estimula sua proliferação e sua diferenciação. O diagrama mostra apenas linfócitos B dando origem a células efetoras secretoras de anticorpo, porém o mesmo princípio aplica-se aos linfócitos T.
A ativação dos linfócitos virgens exige o reconhecimento de complexos peptídeo-MHC apresentados pelas células dendríticas. A natureza do complexo que ativa as células T (i.e., peptídeos ligados a moléculas MHC) assegura que esses linfócitos possam interagir apenas com outras células (visto que as moléculas MHC são proteínas de superfície celular), e não com o antígeno livre. Essa característica é necessária, visto que todas as funções dos linfócitos T dependem de suas interações físicas com outras células. Para responder, as células T precisam reconhecer não apenas os antígenos, mas também outras moléculas, denominadas coestimuladoras, que são induzidas pelos micro-organismos a serem expressas nas superfícies das APC. O reconhecimento do antígeno fornece especificidade à resposta imunológica, e a necessidade de coestimulação assegura que as células T respondam a micro-organismos (os indutores das moléculas coestimuladoras), e não a substâncias inofensivas.
Os linfócitos B utilizam seus receptores de antígenos (moléculas de anticorpos ligadas à membrana) para reconhecer antígenos de várias composições químicas diferentes.
A ocupação dos receptores de antígenos e outros sinais desencadeiam a proliferação e a diferenciação dos linfócitos. As respostas e as funções dos linfócitos T e B diferem de maneira importante, razão pela qual é melhor considerá-las separadamente.
Imunidade Celular: Ativação dos Linfócitos T e Eliminação dos Micro-organismos Intracelulares
Os linfócitos T CD4+ auxiliares ativados proliferam e diferenciam-se em células efetoras cujas funções são mediadas, em grande parte, por citocinas secretadas. Uma das respostas mais iniciais das células T auxiliares CD4+ é a secreção da citocina, a interleucina 2 (IL-2). A IL-2 é um fator de crescimento que atua sobre os linfócitos ativados por antígenos e que estimula a sua proliferação (expansão clonal). Parte da progênie diferencia-se em células efetoras, que podem secretar diferentes conjuntos de citocinas e, assim, desempenhar diferentes funções. Muitas dessas células efetoras deixam os órgãos linfoides em que foram geradas e migram para os locais de infecção e inflamação que a acompanha. Quando esses efetores diferenciados são novamente expostos a micro-organismos associados a células, são ativados para desempenhar as funções responsáveis pela eliminação dos micro-organismos. Algumas células T efetoras da linhagem de células auxiliares CD4+ secretam citocinas que recrutam leucócitos e que estimulam a produção de substâncias microbicidas nos fagócitos. Por conseguinte, essas células T auxiliares ajudam os fagócitos a destruir os patógenos. Outras células T efetoras CD4+ secretam citocinas que estimulam a produção de uma classe especial de anticorpo, denominada imunoglobulina E (IgE), e ativam leucócitos denominados eosinófilos, que são capazes de matar parasitas que podem ser demasiado grandes para serem fagocitados. Conforme será discutido adiante, algumas células T auxiliares CD4+ permanecem nos órgãos linfoides e estimulam as respostas das células B.
Os linfócitos CD8+ ativados proliferam e diferenciam-se em CTL, que destroem as células que contêm micro-organismos no citoplasma. Esses micro-organismos podem ser vírus infectando vários tipos de células, ou bactérias que são ingeridas pelos macrófagos mas que escapam das vesículas fagocíticas para o citoplasma (onde são inacessíveis aos mecanismos de destruição dos fagócitos, que se limita, em grande parte, às vesículas). Ao destruir as células infectadas, os CTL eliminam os reservatórios da infecção.
Imunidade Humoral: Ativação dos Linfócitos B e Eliminação dos Micro-organismos Extracelulares
Uma vez ativados, os linfócitos B proliferam e diferenciam-se em células que secretam diferentes classes de anticorpos, com funções distintas. A resposta das células B a antígenos proteicos exige sinais ativadores (“auxiliares”) das células T CD4+ (que é a razão histórica para a designação dessas células T como células “auxiliares”). As células B podem responder a numerosos antígenos não proteicos sem a participação de outras células.
Parte da progênie dos clones expandidos de células B diferencia-se em plasmócitos secretores de anticorpos. Cada plasmócito secreta anticorpos que têm o mesmo sítio de ligação de antígeno que os anticorpos de superfície celular (receptores das células B) que inicialmente reconheceram o antígeno. Os polissacarídios e os lipídios estimulam a secreção principalmente da classe de anticorpos denominada IgM. Já os antígenos proteicos, após induzirem uma produção inicial de IgM, causam a produção de anticorpos de classes funcionalmente distintas de anticorpos (IgG, IgA ou IgE). A produção desses anticorpos diferentes é denominada mudança de classe e exige a ação das células T auxiliares; esta proporciona plasticidade à resposta dos anticorpos, possibilitando o desempenho de muitas funções. As células T auxiliares também estimulam a produção de anticorpos com afinidade aumentada pelo antígeno. Esse processo, denominado maturação da afinidade, melhora a qualidade da resposta imune humoral.
A resposta imune humoral combate os micro-organismos de muitas maneiras. Os anticorpos ligam-se aos micro-organismos e os impedem de infectar as células, “neutralizando”, assim, os micro-organismos ao bloquear a sua capacidade de infectar as células do hospedeiro ou de colonizar os tecidos. De fato, os anticorpos constituem os únicos mecanismos da imunidade adquirida que impedem o estabelecimento de uma infecção; esta é a razão pela qual um dos objetivos essenciais da vacinação consiste em estimular a produção de anticorpos potentes. Os anticorpos IgG recobrem os micro-organismos e os transformam em alvos para a fagocitose, visto que os fagócitos (neutrófilos e macrófagos) expressam receptores para as caudas de IgG. A IgG e a IgM ativam o sistema complemento através da via clássica, e os produtos do complemento promovem a fagocitose ou a destruição direta dos micro-organismos. Alguns anticorpos desempenham papéis especiais em locais anatômicos específicos. A IgA é secretada pelos epitélios da mucosa e neutraliza os micro-organismos presentes no lúmen dos tratos respiratório e gastrointestinal (e em outros tecidos da mucosa). A IgG materna é transportada ativamente através da placenta e protege o recém-nascido até que o sistema imunológico do lactente amadureça. Os anticorpos têm, em sua maioria, meia-vida de poucos dias; entretanto, alguns anticorpos IgG possuem meia-vida de cerca de 3 semanas. Alguns plasmócitos secretores de anticorpos migram até a medula óssea e vivem durante anos, continuando a produzir baixos níveis de anticorpos. Os anticorpos que são secretados por esses plasmócitos de vida longa fornecem uma proteção imediata caso o micro-organismo volte a infectar o indivíduo. Uma proteção mais efetiva é proporcionada pelas células de memória, que quando ativadas pelo micro-organismo, diferenciam-se rapidamente para produzir grandes números de plasmócitos.
Memória Imunológica
Uma resposta imunológica efetiva elimina os micro-organismos que iniciaram a resposta. Segue-se então uma fase de contração, em que os clones de linfócitos expandidos morrem e a homeostasia é restaurada.
A ativação inicial dos linfócitos gera células de memória de vida longa, que podem sobreviver durante anos após a infecção. As células de memória são mais efetivas no combate aos micro-organismos do que os linfócitos virgens, visto que, conforme foi assinalado anteriormente, as células de memória representam um reservatório expandido de linfócitos específicos para um determinado antígeno (mais numerosos que as células virgens específicas para o antígeno) e respondem mais rapidamente e de modo mais efetivo contra o antígeno do que as células virgens. Esta é a razão pela qual a geração de respostas de memória constitui outra meta importante da vacinação. As propriedades dos linfócitos de memória serão discutidas em capítulos posteriores.
No restante deste livro, descrevemos de modo pormenorizado o reconhecimento, a ativação, a regulação e as fases efetoras das respostas imunes natural e adaptativa. Os princípios apresentados neste capítulo reaparecem ao longo do livro.
• A imunidade protetora contra micro-organismos é mediada pelas reações iniciais da imunidade inata e pelas respostas posteriores da imunidade adaptativa. As respostas imunológicas naturais são estimuladas por estruturas moleculares compartilhadas por grupos de micro-organismos e por moléculas expressas pelas células lesionadas do hospedeiro. A imunidade adquirida é específica para diferentes antígenos microbianos e não microbianos e é aumentada por reexposições ao antígeno (memória imunológica).
• A imunidade humoral é mediada pelos linfócitos B e seus produtos secretados, os anticorpos, e atua na defesa contra micro-organismos extracelulares. A imunidade celular é mediada pelos linfócitos T e seus produtos, como as citocinas, sendo importante na defesa contra micro-organismos intracelulares.
• A imunidade pode ser adquirida por uma resposta ao antígeno (imunidade ativa), ou pela transferência de anticorpos ou de células provenientes de um indivíduo imunizado (imunidade passiva).
• O sistema imune possui várias propriedades, que são de importância fundamental para suas funções normais. Essas propriedades incluem a especificidade para diferentes antígenos, um repertório diverso capaz de reconhecer uma ampla variedade de antígenos, a memória da exposição ao antígeno, a capacidade de rápida expansão de clones de linfócitos específicos para determinado antígeno em resposta ao antígeno, respostas especializadas a diferentes micro-organismos, manutenção da homeostasia e capacidade de discriminar entre antígenos estranhos e antígenos próprios.
• Os linfócitos são as únicas células capazes de reconhecer especificamente antígenos e, portanto, constituem as principais células da imunidade adaptativa. As duas subpopulações principais de linfócitos são as células B e as células T, que diferem nos seus receptores de antígenos e em suas funções. As células apresentadoras de antígenos especializadas capturam antígenos microbianos e os apresentam para reconhecimento pelos linfócitos. A eliminação dos antígenos frequentemente exige a participação de várias células efetoras.
• A resposta imunológica adaptativa é iniciada pelo reconhecimento de antígenos estranhos por linfócitos específicos. Os linfócitos respondem através de sua proliferação e diferenciação em células efetoras, cuja função é eliminar o antígeno, e em células de memória, que apresentam uma resposta intensificada em exposições posteriores ao mesmo antígeno. A ativação dos linfócitos requer a presença do antígeno e sinais adicionais, que podem ser fornecidos pelos micro-organismos ou pelas respostas imunológicas naturais a eles.
• Os linfócitos T CD4+auxiliares ajudam os macrófagos a eliminar os micro-organismos ingeridos e ajudam as células B a produzir anticorpos. Os CTL CD8+ destroem as células que contêm patógenos intracelulares, eliminando, assim, os reservatórios de infecção. Os anticorpos, que são os produtos dos linfócitos B, neutralizam a capacidade infectante dos micro-organismos e promovem a sua eliminação pelos fagócitos e pela ativação do sistema complemento.
Burnet FM. A modification of Jerne’s theory of antibody production using the concept of clonal selection. Australian Journal of Science. 1957;20:67-69.
Flajnik MF, du Pasquier L. Evolution of innate and adaptive immunity: can we draw a line? Trends in Immunology. 2004;25:640-644.
Jerne NK. The natural-selection theory of antibody formation. Proceedings of the National Academy of Sciences U S A. 1955;41:849-857.
Litman GW, Rast JP, Fugmann SD. The origins of vertebrate adaptive immunity. Nature Reviews Immunology. 2010;10:543-553.
Silverstein AM. Paul Ehrlich’s Receptor Immunology: The Magnificent Obsession. New York: Academic Press, 2001.
Silverstein AM. Cellular versus humoral immunology: a century- long dispute. Nature Immunology. 2003;4:425-428.